Торможение в цнс. Центральное торможение по сеченову Явление центрального торможения было открыто
Интегративная и координационная деятельность центральных нервных образований осуществляется при обязательном участии тормозных процессов.
Торможение в центральной нервной системе - активный процесс, проявляющийся внешне в подавлении или в ослаблении процесса возбуждения и характеризующийся определенной интенсивностью и длительностью.
Торможение в норме неразрывно связано с возбуждением, является его производным, сопутствует возбудительному процессу, ограничивая и препятствуя чрезмерному распространению последнего. При этом торможение часто ограничивает возбуждение и вместе с ним формирует сложную мозаику активированных и заторможенных зон в центральных нервных структурах. Формирующий эффект тормозного процесса развивается в пространстве и во времени. Торможение - врожденный процесс, постоянно совершенствующийся в течение индивидуальной жизни организма.
При значительной силе фактора, вызвавшего торможение, оно может распространяться на значительное пространство, вовлекая в тормозной процесс большие популяции нервных клеток.
История развития учения о тормозных процессах в центральной нервной системе начинается с открытия И. М. Сеченовым эффекта центрального торможения (химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные реакции). Вначале предположение о существовании специфических тормозных нейронов, обладающих способностью оказывать тормозные влияния на другие нейроны, с которыми имеются синаптические контакты, диктовалось логической необходимостью для объяснения сложных форм координационной деятельности центральных нервных образований. Впоследствии это предположение нашло прямое экспериментальное подтверждение (Экклс, Реншоу), когда было показано существование специальных вставочных нейронов, имеющих синаптические контакты с двигательными нейронами. Активация этих вставочных нейронов закономерно приводила к торможению двигательных нейронов. В зависимости от нейронного механизма, способа вызывания тормозного процесса в ЦНС различают несколько видов торможения: постсинаптическое, пресинаптическое, пессимальное.
Постсинаптическое торможение - основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной медиатор. Тормозной эффект таких нейронов обусловливается специфической природой медиатора - химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Химическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических потенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД.
Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки. Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мотонейронах, оказывая на них тормозное действие.
Пресинаптическое торможение развертывается в аксоаксональных синапсах, блокируя распространение возбуждения по аксону. Пресинаптическое торможение часто выявляется в структурах мозгового ствола, в спинном мозге
Пессимальное торможение представляет собой вид торможения центральных нейронов. Оно наступает при высокой частоте раздражения. В первый момент возникает высокая частота ответного возбуждения. Через некоторое время стимулируемый центральный нейрон, работая в таком режиме, переходит в состояние торможения.
Опыт Сеченова (сеченовское торможение ). Торможение в центральной нервной системе было открыто И.М.Сеченовым в 1862 г. Он наблюдал возникновение торможения спинномозговых рефлексов при раздражении промежуточного мозга (зрительных бугров) лягушки кристалликом поваренной соли. Внешне это выражалось в значительном уменьшении рефлекторной реакции (увеличении времени рефлекса) или ее прекращении. Снятие кристаллика поваренной соли приводило к восстановлению исходного времени рефлекса.
 |
|
 Б Б
|
Опыт гольца .
Было показано, что торможение может развиваться при встрече двух сильных раздражений (опыт Гольца).
Опыт Гольца: если одну лапку лягушки опустить в кислоту, то лягушка ее отдернет, если одну в кислоту, а другую сильно сжать пинцетом – отдергивания из кислоты не происходит. Если срезать большие полушария и гладить лягушку по голове, то она непременно квакает при каждом поглаживании. Мы имеем, значит, вполне закономерный квакательный рефлекс. Но стоит наряду с поглаживанием раздражать другое место лягушки, например надавить на палец лапы, чтобы этот рефлекс кваканья исчез.
Торможение (физиология)
Торможение - в физиологии - активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения. Обеспечивает (вместе с возбуждением) нормальную деятельность всех органов и организма в целом. Имеет охранительное значение (в первую очередь для нервных клеток коры головного мозга), защищая нервную систему от перевозбуждения.
И. П. Павлов называл иррадиацию торможения по коре больших полушарий головного мозга «проклятым вопросом физиологии».
Центральное торможение
Центральное торможение открыто в 1862 г. И. М. Сеченовым . В процессе опыта он удалил у лягушки головной мозг на уровне зрительных бугров и определял время сгибательного рефлекса. Затем на зрительные бугры помещался кристалл соли в результате чего наблюдалось увеличение продолжительности времени рефлекса. Это наблюдение позволило И. М. Сеченову высказать мнение о явлении торможения в ЦНС. Данный тип торможения называют сеченовским или центральным .
Ухтомский объяснил результаты с позиции доминанты. В зрительных буграх - доминанта возбуждения, которая подавляет действие спинного мозга.
Введенский объяснил результаты с позиции отрицательной индукции. Если в центральной нервной системе возникает возбуждение в определенном нервном центре, то вокруг очага возбуждения индуцируется торможение. Современное объяснение: при раздражении зрительных бугров возбуждается каудальный отдел ретикулярной формации. Эти нейроны возбуждают тормозные клетки спинного мозга (клетки Реншоу ), которые тормозят активность альфа-мотонейронов спинного мозга.
Первичное торможение
Первичное торможение возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы.
Виды первичного торможения
Возвратное - нейрон воздействует на клетку, которая в ответ тормозит этот же нейрон.
Реципрокное - это взаимное торможение, при котором возбуждение одной группы нервных клеток обеспечивает торможение других клеток через вставочный нейрон .
Латеральное - тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клеткамисетчатки , что создает условия для более четкого видения предмета.
Возвратное облегчение - нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками.
Постсинаптическое - основной вид первичного торможения, вызывается возбуждением клеток Реншоу и вставочных нейронов. При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение. Примеры первичного торможения:
Пресинаптическое - возникает в обычных нейронах, связано с процессом возбуждения.
Вторичное торможение
Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
Виды вторичного торможения
Пессимальное торможение - это вторичное торможение, которое развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под действием множественной импульсации.
Торможение вслед за возбуждением возникает в обычных нейронах и также связано с процессом возбуждения. В конце акта возбуждения нейрона в нем может развиваться сильная следовая гиперполяризация. В то же время возбуждающий постсинаптический потенциал не может довести деполяризацию мембраны докритического уровня деполяризации , потенциалзависимые натриевые каналы не открываются и потенциал действия не возникает.
Периферическое торможение
Открыто братьями Вебер в 1845 г. В качестве примера можно привести торможение деятельности сердца (снижение ЧСС ) при раздражении блуждающего нерва .
Условное и безусловное торможение
Термины «условное» и «безусловное» торможение предложены И. П. Павловым.
Условное торможение
Условное, или внутреннее, торможение - форма торможения условного рефлекса, возникающее при неподкреплении условных раздражителей безусловными. Условное торможение является приобретенным свойством и вырабатывается в процессе онтогенеза. Условное торможение является центральным торможением и ослабевает с возрастом.
Безусловное торможение
Безусловное (внешнее) торможение - торможение условного рефлекса, возникающее под действием безусловных рефлексов (например, ориентировочного рефлекса ). И. П. Павлов относил безусловное торможение к врожденным свойствам нервной системы, то есть безусловное торможение является формой центрального торможения.
Торможение
Координирующая функция локальных нейронных сетей помимо усиления может выражаться и в ослаблении слишком интенсивной активности нейронов за счет их торможения.
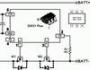
Рис.8.1 .Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга
1 - мотонейрон; 2 - тормозный интернейрон; 3 - афферентные терминали.
Торможение , как особый нервный процесс, характеризуется отсутствием способности к активному распространению по нервной клетке и может быть представлено двумя формами - первичным и вторичным торможением.
Первичное торможение обусловлено наличием специфических тормозных структур и развивается первично без предварительного возбуждения. Примером первичного торможения является так называемое реципрокное торможение мыщц-антагонистов , обнаруженное в спинальных рефлекторых дугах. Суть этого явления состоит в том, что если активируются проприорецепторы мышцы-сгибателя, то они через первичные афференты одновременно возбуждают мотонейрон данной мышцы-сгибателя и через коллатераль афферентного волокна тормозный вставочный нейрон. Возбуждение вставочного нейрона приводит к постсинаптическому торможению мотонейрона антагонистической мышцы-разгибателя, на теле которого аксон тормозного интернейрона формирует специализированные тормозные синапсы. Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
Другим примером первичного торможения является открытое Б. Реншоу возвратное торможение . Оно осуществляется в нейронной цепи, которая состоит из мотонейрона и вставочного тормозного нейрона - клетки Реншоу . Импульсы от возбужденного мотонейрона через отходящие от его аксона возвратные кол-латерали активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона. Это торможение реализуется за счет функции тормозных синапсов, которые клетка Реншоу образует на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, позволяющий стабилизировать частоту разрядов моторной клетки и подавить идущую к мышцам избыточную импульсацию.
В ряде случаев клетки Реншоу формируют тормозные синапсы не только на активирующих их мотонейронах, но и на соседних мотонейронах со сходными функциями. Осуществляемое через эту систему торможение окружающих клеток называется латеральным .
Торможение по принципу отрицательной обратной связи встречается не только на выходе, но и на входе моторных центров спинного мозга. Явление подобного рода описано в моносинаптических соединенях афферентных волокон со спинальными мотонейронами, торможение которых при данной ситуации не связано с изменениями в постсинаптической мембране. Последнее обстоятельство позволило определить данную форму торможения как пресинаптическое . Оно обусловлено наличием вставочных тормозных нейронов, к которым подходят коллатерали афферентных волокон. В свою очередь, вставочные нейроны формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению к мотонейронам. В случае избыточного притока сенсорной информации с периферии происходит активация тормозных интернейронов, которые через аксо-аксональные синапсы вызывают деполяризацию афферентных термина-леи и, таким образом, уменьшают количество выделяемого из них медиатора, а следовательно, и эффективность синаптической передачи. Электрофизиологическим показателем этого процесса является снижение амплитуды регистрируемых от мотонейрона ВПСП. Вместе с тем никаких признаков изменений ионной проницаемости или генерации ТПСП в мотонейронах не наблюдается.
Вопрос о механизмах пресинаптического торможения является довольно сложным. По-видимому, медиатором в тормозном аксо-аксональном синапсе является гамма-аминомасляная кислота, которая вызывает деполяризацию афферентных терминалей за счет увеличения проницаемости их мембраны для ионов С1-. Деполяризация снижает амплитуду потенциалов действия в афферентных волокнах и тем самым уменьшает квантовый выброс медиатора в синапсе. Другой возможной причиной деполяризации терминалей может быть повышение наружной концентрации ионов К+ при длительной активации афферентных входов. Следует отметить, что феномен пресинаптического торможения обнаружен не только в спинном мозгу, но и в других отделах ЦНС.
Исследуя координирующую роль торможения в локальных нейронных цепях, следует упомянуть еще об одной форме торможения - вторичном торможении , которое возникает без участия специализированных тормозных структур как следствие избыточной активации возбуждающих входов нейрона. В специальной литературе эту форму торможения определяют как торможение Введенского , который открыл его в 1886 г. при исследовании нервно-мышечного синапса.
Торможение Введенского играет предохранительную роль и возникает при чрезмерной активации центральных нейронов в полисинаптических рефлекторных дугах. Оно выражается в стойкой деполяризации клеточной мембраны, превышающей критический уровень и вызывающей инактивацию Na-каналов, ответственных за генерацию потенциалов действия. Таким образом, процессы торможения в локальных нейронных сетях уменьшают избыточную активность и участвуют в поддержании оптимальных режимов импульсной активности нервных клеток.
ТОРМОЖЕНИЕ В ЦНС. ВИДЫ И ЗНАЧЕНИЕ.
Проявление и осуществление рефлекса возможно только при ограничении распространения возбуждения с одних нервных центров на другие. Это достигается взаимодействием возбуждения с другим нервным процессом, противоположным по эффекту процессом торможения.
Почти до середины XIX века физиологи изучали и знали только один нервный процесс - возбуждение.
Явления торможения в нервных центрах, т.е. в центральной нервной системе были впервые открыты в 1862 году И.М.Сеченовым ("сеченовское торможение”). Это открытие сыграло в физиологии не меньшую роль, чем сама формулировка понятия рефлекса, так как торможение обязательно участвует во всех без исключения нервных актах. И.М.Сеченов обнаружил явление центрального торможения при раздражении промежуточного мозга теплокровных. В 1880 году немецкий физиолог Ф.Гольц установил торможение спинальных рефлексов. Н.Е. Введенский в результате серий опытов по парабиозу вскрыл интимную связь процессов возбуждения и торможения и доказал, что природа этих процессов едина.
Торможение - местный нервный процесс, приводящий к угнетению или предупреждению возбуждения. Торможение является активным нервным процессом, результатом которого служит ограничение или задержка возбуждения. Одна из характерных черт тормозного процесса- отсутствие способности к активному распространению по нервным структурам.
В настоящее время в центральной нервной системе выделяют два вида торможения:торможение центральное (первичное), являющееся результатом возбуждения (активации) специальных тормозных нейронов иторможение вторичное, которое осуществляется без участия специальных тормозных структур в тех самых нейронах в которых происходит возбуждение.
Центральное торможение(первичное) - нервный процесс, возникающий в ЦНС и приводящий к ослаблению или предотвращению возбуждения. Согласно современным представлениям центральное торможение связано с действием тормозных нейронов или синапсов, продуцирующих тормозные медиаторы (глицин, гаммааминомасляную кислоту), которые вызывают на постсинаптической мембране особый тип электрических изменений, названных тормозными постсинаптическими потенциалами (ТПСП) или деполяризацию пресинаптического нервного окончания, с которым контактирует другое нервное окончание аксона. Поэтому выделяют центральное (первичное) постсинаптическое торможение и центральное (первичное) пресинаптическое торможение.
Постсинаптическое торможение (лат. post позади, после чего-либо + греч. sinapsis соприкосновение, соединение) - нервный процесс, обусловленный действием на постсинаптическую мембрану специфических тормозных медиаторов (глицин, гаммааминомаслянная кислота), выделяемых специализированными пресинаптическими нервными окончаниями. Медиатор, выделяемый ими, изменяет свойства постсинаптической мембраны, что вызывает подавление способности клетки генерировать возбуждение. При этом происходит кратковременное повышение проницаемости постсинаптической мембраны к ионам К+ или CI, вызывающее снижение ее входного электрического сопротивления и генерацию тормозного постсинаптического потенциала (ТПСП). Возникновение ТПСП в ответ на афферентное раздражение обязательно связано с включением в тормозной процесс дополнительного звена - тормозного интернейрона, аксональные окончания которого выделяют тормозной медиатор. Специфика тормозных постсинаптических эффектов впервые была изучена на мотонейронах млекопитающих (Д. Экклс, 1951). В дальнейшем первичные ТПСП были зарегистрированы в промежуточных нейронах спинного и продолговатого мозга, в нейронах ретикулярной формации, коры больших полушарий, мозжечка и таламических ядер теплокровных животных.
Известно, что при возбуждении центра сгибателей одной из конечностей центр ее разгибателей тормозится и наоборот. Д. Экклс выяснил механизм этого явления в следующем опыте. Он раздражал афферентный нерв, вызывающий возбуждение мотонейрона, иннервирующего мышцу - разгибатель.
Нервные импульсы, дойдя до афферентного нейрона в спинномозговом ганглии, направляются по его аксону в спинном мозге по двум путям: к мотонейрону, иннервирующему мышцу - разгибатель, возбуждая ее и по коллатерам к промежуточному тормозному нейрону, аксон которого контактирует с мотонейроном иннервирующим мышцу - сгибатель, вызывая таким образом торможение антагонистической мышцы. Этот вид торможения был обнаружении в промежуточных нейронах всех уровней центральной нервной системы при взаимодействии антагонистических центров. Он был назван поступательным постсинаптическим торможением . Этот вид торможения координирует, распределяет процессы возбуждения и торможения между нервными центрами.
Возвратное (антидромное) постсинаптическое торможение (греч. antidromeo бежать в противоположном направлении) - процесс регуляции нервными клетками интенсивности поступающих к ним сигналов по принципу отрицательной обратной связи. Он заключается в том, что коллатерали аксонов нервной клетки устанавливают синаптические контакты со специальными вставочными нейронами (клетки Реншоу), роль которых заключается в воздействии на нейроны, конвергирующие на клетке, посылающей эти аксонные коллатерали (рис. 87). По такому принципу осуществляетсяторможение мотонейронов.
Возникновение импульса в мотонейроне млекопитающих не только активирует мышечные волокна, но через коллатерали аксона активирует тормозные клетки Реншоу. Последние устанавливают синаптические связи с мотонейронами. Поэтому усиление импульсации мотонейрона ведет к большей активации клеток Реншоу, вызывающей усиление торможения мотонейронов и уменьшение частоты их импульсации. Термин "антидромное” употребляется потому, что тормозной эффект легко вызывается антидромными импульсами, рефлекторно возникающими в мотонейронах.
Чем сильнее возбужден мотонейрон, чем больше сильные импульсы идут к скелетным мышцам по его аксону, тем интенсивнее возбуждается клетка Реншоу, которая подавляет активность мотонейрона. Следовательно, в нервной системе существует механизм, оберегающий нейроны от чрезмерного возбуждения. Характерная особенность постсинаптического торможения заключается в том, что оно подавляется стрихнином и столбнячным токсином (на процессы возбуждения эти фармакологические вещества не действуют).
В результате подавления постсинаптического торможения нарушается регуляция возбуждения в цнс, возбуждение разливается ("диффундирует”) по всей цнс, вызывая перевозбуждение мотонейронов и судорожные сокращения групп мышц (судороги).
Торможение ретикулярное (лат. reticularis - сетчатый) - нервный процесс развивающийся в спинальных нейронах под влиянием нисходящей импульсации из ретикулярной формации (гигантское ретикулярное ядро продолговатого мозга). Эффекты, создаваемые ретикулярными влияниями, по функциональному действию сходны с возвратным торможением, развивающимся на мотонейронах. Влияние ретикулярной формации вызывают стойкие ТПСП, охватывающие все мотонейроны независимо от их функциональной принадлежности. В этом случае, так же как и при возвратном торможении мотонейронов происходит ограничение их активности. Между таким нисходящим контролем со стороны ретикулярной формации и системочй возвратного торможения через клетки Реншоу существует определенное взаимодействие, и клетки Реншоу находятся под постоянным тормозящем контролем со стороны двух структур. Тормозящее влияние со стороны ретикулярной формации являются дополнительным фактором в регуляции уровня активности мотонейронов.
Первичное торможение может вызываться механизмами иной природы, не связанными с изменениями свойств постсинаптической мембраны. Торможение в этом случае возникает на пресинаптической мембране (синаптическое и пресинаптическое торможение).
Синаптическое торможение (греч. sunapsis соприкосновение, соединение) - нервный процесс, основанный на взаимодействии медиатора, секретируемого и выделяемого пресинаптическими нервными окончаниями, со специфическими молекулами постсинаптической мембраны. Возбуждающий или тормозной характер действия медиатора зависит от природы каналов, которые открываются в постсинаптической мембране. Прямое доказательство наличия в цнс специфических тормозящих синапсов было впервые получено Д. Ллойдом (1941).
Данные относительно электрофизиологических проявлений синаптического торможения: наличие синаптической задержки, отсутствие электрического поля в области синаптических окончаний дали основание считать его следствием химического действия особого тормозящего медиатора, выделяемого синаптическими окончаниями. Д. Ллойд показал, что если клетка находится в состоянии деполяризации, то тормозной медиатор вызывает гиперполяризацию, в то время как на фоне гиперполяризации постсинаптической мембраны он вызывает ее деполяризацию.
Пресинаптическое торможение (лат. praе -впереди чего-либо + греч. sunapsis соприкосновение, соединение) - частный случай синаптических тормозных процессов, проявляющихся в подавлении активности нейрона в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким-либо изменениям. Пресинаптическое торможение осуществляется посредством специальных тормозных интернейронов. Его структурной основой являются аксо-аксональные синапсы, образованные терминалиями аксонов тормозных интернейронов и аксональными окончаниями возбуждающих нейронов.
При этом окончание аксона тормозного нейрона является пресимпатическим по отношению к терминали возбуждающего нейрона, которая оказывается постсинаптической по отношению к тормозному окончанию и пресинаптической по отношению к активируемой им нервной клетки. В окончаниях пресинаптического тормозного аксона освобождается медиатор, который вызывает деполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембраны для CI. Деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание аксона. В результате происходит угнетение процесса высвобождения медиатора возбуждающими нервными окончаниями и снижение амплитуды возбуждающего постсинаптического потенциала.
Характерной особенностью пресинаптической деполяризации является замедленное развитие и большая длительность (несколько сотен миллисекунд), даже после одиночного афферентного импульса.
Пресинаптическое торможение существенно отличается от постсинаптического и в фармакологическом отношении. Стрихнин и столбнячный токсин не влияют на его течение. Однако наркотизирующие вещества (хлоралоза, нембутал) значительно усиливают и удлиняют пресинаптическое торможение. Этот вид торможения обнаружен в различных отделах цнс. Наиболее часто оно выявляется в структурах мозгового ствола и спинного мозга. В первых исследованиях механизмов пресинаптического торможения считалось, что тормозное действие осуществляется в точке, отдаленной от сомы нейрона, поэтому его называли "отдаленным” торможением.
Функциональное значение пресинаптического торможения, охватывающего пресинаптические терминали, по которым поступают афферентные импульсы, заключается в ограничении поступления к нервным центрам афферентной импульсации. Пресинаптическое торможение в первую очередь блокирует слабые асинхронные афферентные сигналы и пропускает более сильные, следовательно, оно служит механизмом выделения, вычленения более интенсивных афферентных импульсов из общего потока. Это имеет огромное приспособительное значение для организма, так как из всех афферентных сигналов, идущих к нервным центрам, выделяются самые главные, самые необходимые для данного конкретного времени. Благодаря этому нервные центры, нервная система в целом освобождается от переработки менее существенной информации.
Вторичное торможение - торможение осуществляющееся теми же нервными структурами, в которых происходит возбуждение. Этот нервный процесс подробно изложен в работах Н.Е. Введенского (1886, 1901г.г.).
Торможение реципрокное (лат. reciprocus - взаимный) - нервный процесс, основанный на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через посредство вставочных нейронов торможение других групп клеток. Реципрокные отношения возбуждения и торможения в цнс были открыты и продемонстрированы Н.Е. Введенским: раздражение кожи на задней лапке у лягушки вызывает ее сгибание и торможение сгибания или разгибания на противоположной стороне. Взаимодействие возбуждения и торможения является общим свойством всей нервной системы и обнаруживается как в головном, так и в спинном мозге. Экспериментально доказано, что нормальное выполнение каждого естественного двигательного акта основано на взаимодействии возбуждения и торможения на одних и тех же нейронах цнс.
Общее центральное торможение - нервный процесс, развивающийся при любой рефлекторной деятельности и захватывавающий почти всю цнс, включая центры головного мозга. Общее центральное торможение обычно проявляется раньше возникновения какой-либо двигательной реакции. Оно может проявляться при такой малой силе раздражения при которой двигательный эффект отсутствует. Такого вида торможение было впервые описано И.С. Беритовым (1937). Оно обеспечивает концентрацию возбуждения других рефлекторных или поведенческих актов, которые могли бы возникнуть под влиянием раздражений. Важная роль в создании общего центрального торможения принадлежит желатинозной субстанции спинного мозга.
При электрическом раздражении желатинозной субстанции у спинального препарата кошки происходит общее торможение рефлекторных реакций, вызываемых раздражением сенсорных нервов. Общее торможение является важным фактором в создании целостной поведенческой деятельности животных, а также в обеспечении избирательного возбуждения определенных рабочих органов.
Парабиотическое торможение развивается при патологических состояниях, когда лабильность структур центральной нервной системы снижается или происходит очень массивное одновременное возбуждение большого числа афферентных путей, как, например, при травматическом шоке.
Некоторые исследователи выделяют еще один вид торможения - торможение вслед за возбуждением . Оно развивается в нейронах после окончания возбуждения в результате сильной следовой гиперполяризации мембраны (постсинаптической).
Торможение – активный процесс, возникающий при действии раздражителей на ткань, проявляется в подавлении другого возбуждения, функционального отправления ткани нет.
Торможение может развиваться только в форме локального ответа.
Выделяют два типа торможения :
1) первичное . Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения под воздействием тормозного медиатора. Различают два вида первичного торможения:
пресинаптическое в аксо-аксональном синапсе;
постсинаптическое в аксодендрическом синапсе.
2) вторичное . Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. Виды вторичного торможения:
запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона;
пессимальное, возникающее при высокой частоте раздражения; парабиотическое, возникающее при сильно и длительно действующем раздражении;
торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения;
торможение по принципу отрицательной индукции;
торможение условных рефлексов.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выраженными. Возбуждение непременно сменяется торможением, и наоборот, т. е. между торможением и возбуждением существуют индукционные отношения.
Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Торможение в ЦНС может возникать при одновременном поступлении в спинной мозг нервных импульсов различной силы с нескольких раздражителей. Более сильное раздражение тормозит рефлексы, которые должны были наступать в ответ на более слабые.
В 1862 г. И. М. Сеченов открыл явление центрального торможения . Он доказал в своем опыте, что раздражение кристалликом хлорида натрия зрительных бугров лягушки (большие полушария головного мозга удалены) вызывает торможение рефлексов спинного мозга. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась. Результат этого опыта позволил И. М. Сеченому сделать заключение, что в ЦНС наряду с процессом возбуждения развивается процесс торможения, который способен угнетать рефлекторные акты организма. Н. Е. Введенский высказал предположение, что в основе явления торможения лежит принцип отрицательной индукции: более возбудимый участок в ЦНС тормозит активность менее возбудимых участков.
Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную формацию ствола мозга): возбуждение ретикулярной формации повышает активность тормозных нейронов спинного мозга – клеток Реншоу, что приводит к торможению α-мотонейронов спинного мозга и угнетает рефлекторную деятельность спинного мозга.
Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, ГАМК и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП ) и, как следствие, - снижение возбудимости нейрона вплоть до полной его рефрактерности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью.
Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Чтобы понять, как тормозные синапсы тормозят постсинаптический нейрон, нужно вспомнить, что мы знаем о потенциале Нернста для ионов Сl-. Мы рассчитали, что он равен примерно -70 мВ. Этот потенциал отрицательнее, чем мембранный потенциал покоя нейрона, равный -65 мВ. Следовательно, открытие хлорных каналов будет способствовать движению отрицательно заряженных ионов Сl- из внеклеточной жидкости внутрь. Это сдвигает мембранный потенциал в направлении более отрицательных значений по сравнению с покоем приблизительно до уровня -70 мВ.
Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией . Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют ТПСП .
ВНС обеспечивает экстраорганную и внутриорганную регуляцию функций организма и включает в себя три компонента:1)симпатический;2)парасимпатический;3)метсимпатический.
Вегетативная нервная система обладает рядом анатомических и физиологических особенностей, которые определяют механизмы ее работы.
Анатомические свойства :
1. Трехкомпонентное очаговое расположение нервных центров. Низший уровень симпатического отдела представлен боковыми рогами с VII шейного по III–IV поясничные позвонки, а парасимпатического – крестцовыми сегментами и стволом мозга. Высшие подкорковые центры находятся на границе ядер гипоталамуса (симпатический отдел – задняя группа, а парасимпатический – передняя). Корковый уровень лежит в области шестого-восьмого полей Бродмана (мотосенсорная зона), в которых достигается точечная локализация поступающих нервных импульсов. За счет наличия такой структуры вегетативной нервной системы работа внутренних органов не доходит до порога нашего сознания.
2. Наличие вегетативных ганглиев. В симпатическом отделе они расположены либо по обеим сторонам вдоль позвоночника, либо входят в состав сплетений. Таким образом, дуга имеет короткий преганглионарный и длинный постганглионарный путь. Нейроны пара-симпатического отдела находятся вблизи рабочего органа или в его стенке, поэтому дуга имеет длинный преганглионарный и короткий постганглионарный путь.
3. Эффеторные волокна относятся к группе В и С.
Физиологические свойства:
1. Особенности функционирования вегетативных ганглиев. Наличие феномена мультипликации (одновременного протекания двух противоположных процессов – дивергенции и конвергенции). Дивергенция – расхождение нервных импульсов от тела одного нейрона на несколько постганглионарных волокон другого. Конвергенция – схождение на теле каждого постганглионарного нейрона импульсов от нескольких преганглионарных. Это обеспечивает надежность передачи информации из ЦНС на рабочий орган. Увеличение продолжительности постсинаптического потенциала, наличие следовой гиперполяризации и синоптической задержки способствуют передаче возбуждения со скоростью 1,5–3,0 м/с. Однако импульсы частично гасятся или полностью блокируются в вегетативных ганглиях. Таким образом они регулируют поток информации из ЦНС. За счет этого свойства их называют вынесенными на периферию нервными центрами, а вегетативную нервную систему – автономной.
2. Особенности нервных волокон. Преганглионарные нервные волокна относятся к группе В и проводят возбуждение со скоростью 3-18 м/с, постганглионарные – к группе С. Они проводят возбуждение со скоростью 0,5–3,0 м/с. Так как эфферентный путь симпатического отдела представлен преганглионарными волокнами, а парасимпатического – постганглионарными, то скорость передачи импульсов выше у парасимпатической нервной системы.
Таким образом, вегетативная нервная система функционирует неодинаково, ее работа зависит от особенностей ганглиев и строения волокон.
Симпатическая нервная система осуществляет иннервацию всех органов и тканей (стимулирует работу сердца, увеличивает просвет дыхательных путей, тормозит секреторную, моторную и всасывательную активность желудочно-кишечного тракта и т. д.). Она выполняет гомеостатическую и адаптационно-трофическую функции.
Ее гомеостатическая роль заключается в поддержании постоянства внутренней среды организма в активном состоянии, т. е.симпатическая нервная система включается в работу только при физических нагрузках, эмоциональных реакциях, стрессах, болевых воздействий, кровопотерях.
Адаптационно-трофическая функция направлена на регуляцию интенсивности обменных процессов. Это обеспечивает приспособление организма к меняющимся условиям среды существования.
Таким образом, симпатический отдел начинает действовать в активном состоянии и обеспечивает работу органов и тканей.
Парасимпатическая нервная система является антагонистом симпатической и выполняет гомеостатическую и защитную функции, регулирует опорожнение полых органов.
Гомеостатическая роль носит восстановительный характер и действует в состоянии покоя. Это проявляется в виде уменьшения частоты и силы сердечных сокращений, стимуляции деятельности желудочно-кишечного тракта при уменьшении уровня глюкозы в крови и т. д.
Все защитные рефлексы избавляют организм от чужеродных частиц. Например, кашель очищает горло, чиханье освобождает носовые ходы, рвота приводит к удалению пищи и т. д.
Опорожнение полых органов происходит при повышении тонуса гладких мышц, входящих в состав стенки. Это приводит к поступлению нервных импульсов в ЦНС, где они обрабатывают и по эффекторному пути направляются до сфинктеров, вызывая их расслабление.
Метсимпатическая нервная система представляет собой совокупность микроганглиев, расположенных в ткани органов. Они состоят из трех видов нервных клеток – афферентных, эфферентных и вставочных, поэтому выполняют следующие функции:
20.Функциональные особенности соматической и вегетативной нервной системы. Сравнительная характеристика симпатического, парасимпатического и метасимпатического отделов вегетативной нервной системы.
Первое и основное отличие строения ВНС от строения соматической состоит в расположении эфферентного (моторного) нейрона. В СНС вставочный и моторный нейроны располагаются в сером веществе СМ, в ВНС эффекторный нейрон вынесен на периферию, за пределы СМ, и лежит в одном из ганглиев - пара-, превертебральном или интраорганном. Более того, в метасимпатической части ВНС весь рефлекторный аппарат полностью находится в интрамуральных ганглиях и нервных сплетениях внутренних органов.
Второе отличие касается выхода нервных волокон из ЦНС. Соматические НВ покидают СМ сегментарно и перекрывают иннервацией не менее трех смежных сегментов. Волокна же ВНС выходят из трех участков ЦНС (ГМ, грудопоясничного и крестцового отделов СМ). Они иннервируют все органы и ткани без исключения. Большинство висцеральных систем имеет тройную (симпатическую, пара- и метасимпатическую) иннервацию.
Третье отличие касается иннервации органов соматической и ВНС. Перерезка у животных вентральных корешков СМ сопровождается полным перерождением всех соматических эфферентных волокон. Она не затрагивает дуги автономного рефлекса ввиду того, что ее эффекторный нейрон вынесен в пара- или превертебральный ганглий. В этих условиях эффекторный орган управляется импульсами данного нейрона. Именно это обстоятельство подчеркивает относительную автономию указанного отдела НС.
Четвертое отличие относится к свойствам нервных волокон. В ВНС они в большинстве своем безмякотные или тонкие мякотные, как, например, преганглионарные волокна, диаметр которых не превышает 5 мкм. Такие волокна принадлежат к типу В. Постганглионарные волокна еще тоньше, большая часть их лишена миелиновой оболочки, они относятся к типу С. В отличие от них соматические эфферентные волокна толстые, мякотные, диаметр их составляет 12-14 мкм. Кроме того, пре- и постганглионарные волокна отличаются низкой возбудимостью. Для вызова в них ответной реакции необходима значительно большая, чем для моторных соматических волокон, сила раздражения. Волокна ВНС характеризуются большим рефрактерным периодом и большой хронаксией. Скорость распространения по ним НИ невелика и составляет в преганглионарных волокнах до 18 м/с, в постганглионарных - до 3 м/с. Потенциалы действия волокон ВНС характеризуются большей, чем в соматических эфферентах, длительностью. Их возникновение в преганглионарных волокнах сопровождается продолжительным следовым положительным потенциалом, в постганглионарных волокнах - следовым отрицательным потенциалом с последующей продолжительной следовой гиперполяризацией (300-400 мс).
обеспечивает внутриорганную иннервацию;
являются промежуточным звеном между тканью и экстраорганной нервной системой. При действии слабого раздражителя активируется метсимпатический отдел, и все решается на местном уровне. При поступлении сильных импульсов они передаются через парасимпатический и симпатический отделы к центральным ганглиям, где происходит их обработка.
Метсимпатическая нервная система регулирует работу гладких мышц, входящих в состав большинства органов желудочно-кишечного тракта, миокарда, секреторную активность, местные иммунологические реакции и др.
1. Основная часть
1.1 Биография Ивана Михайловича Сеченова
1.2 Открытия и научные труды И.М. Сеченова
1.3 Влияние трудов И.М. Сеченова на последующее развитие физиологии
1.4 «Рефлексы головного мозга». Основной труд И.М. Сеченова
Заключение
Введение
Иван Михайлович Сеченов (1829-1905) - российский ученый и мыслитель-материалист, создатель физиологической школы, член-корреспондент (1869), почетный член (1904) Петербургской АН.
В классическом труде «Рефлексы головного мозга» (1866) Иван Сеченов обосновал рефлекторную природу сознательной и бессознательной деятельности, показал, что в основе психических явлений лежат физиологические процессы, которые могут быть изучены объективными методами. Открыл явления центрального торможения, суммации в нервной системе, установил наличие ритмических биоэлектрических процессов в центральной нервной системе, обосновал значение процессов обмена веществ в осуществлении возбуждения.
Сеченов так же исследовал и обосновал дыхательную функцию крови. Создатель объективной теории поведения, заложил основы физиологии труда, возрастной, сравнительной и эволюционной физиологии. Труды Сеченова оказали большое влияние на развитие естествознания и теории познания.
Вклад этого ученого в науку метко охарактеризовал Иван Петрович Павлов, назвавший Сеченова «отцом русской физиологии». Действительно, с его именем физиология не только вошла в мировую науку, но и заняла в ней одно из ведущих мест.
Цель работы выявить вклад, внесенный в развитие физиологии человека и животных И.М. Сеченовым.
Задачами для достижения цели являются:
Ознакомиться с биографией И.М. Сеченова;
Рассмотреть работы в области физиологии И.М. Сеченова;
Оценить вклад И.М. Сеченова в физиологию человека и животных, как науку
Основная часть
1 Биография Ивана Михайловича Сеченова
Родился 13 августа 1829 г. в селе Теплый Стан Симбирской губернии (ныне село Сеченово в Нижегородской области). Сын помещика и его бывшей крепостной.
Окончил в 1848 г. Главное инженерное училище в Петербурге. Проходил военную службу в Киеве, вышел в отставку в 1850 г. и через год поступил в Московский университет на медицинский факультет, который окончил с отличием в 1856 г.
Во время стажировки в Германии сблизился с С. П. Боткиным, Д. И. Менделеевым, композитором А. П. Бородиным, художником А. А. Ивановым. Личность Сеченова оказала такое влияние на российскую художественную интеллигенцию того времени, что Н. Г. Чернышевский списывал с него своего Кирсанова в романе «Что делать?», а И. С. Тургенев - Базарова («Отцы и дети»).
В 1860 г. возвратился в Петербург, защитил диссертацию на степень доктора медицинских наук и возглавил кафедру в Медико-хирургической академии, а также лабораторию, где выполнялись исследования в области физиологии, токсикологии, фармакологии, клинической медицины.
С 1876 по 1901 г. преподавал в Московском университете. Более 20 лет жизни Сеченов посвятил изучению газов и дыхательной функции крови, однако наиболее фундаментальными его работами являются исследования рефлексов головного мозга. Именно он открыл феномен центрального торможения, названный сеченовским торможением (1863 г.). Тогда же по предложению Н. А. Некрасова Сеченов написал для журнала «Современник» статью «Попытка ввести физиологические основы в психические процессы», которую цензура не пропустила за «пропаганду материализма». Эта работа под названием «Рефлексы головного мозга» вышла в «Медицинском вестнике» (1866 г.).
В 90-х гг. Сеченов обратился к проблемам психофизиологии и теории познания. Курс лекций, прочитанный им в Московском университете, лёг в основу «Физиологии нервных центров» (1891 г.), где рассматривается широкий спектр нервных явлений - от бессознательных реакций у животных до высших форм восприятия у человека. Затем учёный начал исследования в новой области - физиологии труда.
В 1901 г. Сеченов вышел в отставку. Его имя присвоено 1-й Московской медицинской академии, Институту эволюционной физиологии и биохимии РАН. Академией наук учреждена премия имени Сеченова, присуждаемая раз в три года за выдающиеся исследования по физиологии.
2 Открытия и научные труды И.М. Сеченова
Исследования и сочинения И.М. Сеченова были посвящены в основном терм проблемам: физиологии нервной системы, химизму дыхания и физиологическим основам психической деятельности. Своими работами И.М. Сеченов положил начало отечественной физиологии и создал школу русских физиологов, которая сыграла важную роль в развитии физиологии, психологии и медицины не только в России, но и во всем мире. Его работы по физиологии дыхания крови, газообмену, растворению газов в жидкостях и обмену энергии заложили основы будущей авиационной и космической физиологии.
Диссертация Сеченова стала первым в истории фундаментальным исследованием влияния алкоголя на организм. Необходимо обратить внимания на сформулированные в ней общефизиологические положения и выводы: во-первых, «все движения, носящие в физиологии название произвольных, суть в строгом смысле рефлективные»; во-вторых, «самый общий характер нормальной деятельности головного мозга (поскольку она выражается движением) есть несоответствие между возбуждением и вызываемым им действием - движением»; И наконец, «рефлекторная деятельность головного мозга обширнее, чем спинного».
Сеченов первый осуществил полное извлечение всех газов их крови и определил их количество ы сыворотке и эритроцитах. Особенно важные результаты получены И.М. Сеченовым при изучении роли эритроцитов в переносе и обмене углекислоты. Им впервые было показано, что углекислота находится в эритроцитах не только в состоянии физического растворения и в виде бикарбоната, но и в состоянии нестойкого химического соединения с гемоглобином. На этом основании И.М. Сеченов пришел к выводу, что эритроциты являются переносчиками кислорода от легких к тканям и углекислоты - от тканей к легким.
Вместе с Мечниковым Сеченов обнаружил тормозящие действие блуждающего нерва на сердце черепахи. Оказалось, что при сильном раздражении чувствительных нервов возникают активные двигательные рефлексы, которые вскоре сменяются полным угнетением рефлекторной деятельности. Эту закономерность крупнейший физиолог Н.Е. Введенский, ученик Сеченова, предложил назвать рефлексом Сеченова.
В чрезвычайно тонких опытах Сеченов производил у лягушек четыре разреза мозга и затем наблюдал, как изменяются рефлекторные движения под влиянием каждого их них. Опыты дали любопытные результаты: угнетение отраженной деятельности наблюдалось лишь после разрезов мозга непосредственно перед зрительными буграми и в них самих. Подводя итог первых опытов - с разрезами мозга, Сеченов высказал мысль о существовании в мозге центров, задерживающих отраженные движения: у лягушки они находятся в зрительных буграх.
Так началась вторая серия экспериментов, во время которых Сеченов производил химическое раздражение различных частей мозга лягушки поваренной солью. Выяснилось, что соль, приложенная к поперечному разрезу мозга в ромбическом пространстве, всегда вызывала столь же сильное угнетение отражательной деятельности, как и разрез мозга в этом месте. Угнетение, но не столь сильное наблюдалось и при раздражении поперечного разреза мозга позади зрительных бугров. Такие же результаты дало и электрическое раздражение поперечных разрезов мозга.
Итак, можно сделать выводы. Во-первых, у лягушки механизмы, задерживающие отраженные движения, лежат в зрительных буграх и продолговатом мозге. Во-вторых, механизмы эти следует рассматривать как нервные центры. В-третьих, один из физиологических путей возбуждения этих механизмов к деятельности представляют волокна чувствительных нервов.
Эти эксперименты Сеченова увенчались открытием центрального торможения - особой физиологической функции головного мозга. Тормозной центр в таломической области получил название сеченовского центра.
Открытие процесса торможения было по заслугам оценено еще его современниками. А вот открытие, которое он так же сделал в ходе экспериментов с лягушкой, ретикулоспинальных влияний (влияний ретикулярной формации мозгового ствола на спинномозговые рефлексы) -получило широкое признание лишь начина с 40-ч годов XX века, после выяснения функции ретикулярной формации головного мозга.
К времени 1860-х годов относится еще одно открытие русского ученого. Он доказал, что нервные центры обладают способностью «суммировать чувствительные, поодиночке не действительные, раздражения до импульса, дающего движение, если эти раздражения достаточно часто следуют друг за другом». Явление суммации - важная характеристика нервной деятельности, впервые открытая И.М. Сеченовым в экспериментах на лягушках, было затем установлено и в опытах на других животных, позвоночных и беспозвоночных, и получило универсальное значение.
Наблюдая за поведение и формированием ребенка, Сеченов показал, как врождённые рефлексы усложняются с возрастом, вступают в связи друг с другом и создают всю сложность человеческого поведения. Он описал, что все акты сознательной и бессознательной жизни по способу происхождения - рефлексы.
Сеченов говорил, что рефлекс лежит и в основе и памяти. Это означает, что все произвольные (сознательные) действия в строгом смысле отраженные, т.е. рефлекторные. Следовательно, умение группировать движения человек приобретает повторением соединяющихся рефлексов. В 1866г. Вышло руководство « Физиология нервных центров» , в котором Сеченов обобщил свой опыт.
Осенью 1889 г. В Московском университете ученый прочел курс лекций по физиологии, который стал основой обобщающего труда «Физиологии нервных центров» (1891 г.). В этой работе был осуществлен анализ различных нервных явлений - от бессознательных реакций у спинальных животных до высших форм восприятия у человека. В 1894г. Он публикует «Физиологические критерии для установки длины рабочего дня», а в 1901 г. - «Очерк рабочих движений человека».
И.М. Сеченов является одним из основоположников отечественной электрофизиологии. Его монография « О животном электричестве» (1862 г.) была первой работой по электрофизиологии в России.
С именем Сеченова связано создание первой в России физиологической научной школы, которая сформировалась и развивалась в Медико-хирургической академии, Новороссийском, Петербургском и Московском университетах. В медико-хирургической Академии Ивам Михайлович ввел в лекционную практику метод демонстрации эксперимента. Это способствовало тесной связи педагогического процесса с исследовательской работой и в значительной степени предопределило успех Сеченова на пути научной школы.
Открытия И.М.Сеченова неопровержимо доказали, что психическая деятельность, как и телесная, подчинена вполне определенным объективным законам, обусловлена естественными материальными причинами, а представляет собой проявление какой-то особой, независимой от тела от окружающих условий «души. Таким образом, был положен конец религиозно-идеалистическому обособлению психического от физического и заложены основы для научного материалистического понимания душевной жизни человека. И.М. Сеченов доказал, что первая причина всякого человеческого действия, поступка, коренится не во внутреннем мире человека, а вне его, в конкретных условиях его жизни и деятельности, и что без внешнего чувственного возбуждения никакая мысль невозможна. Этим И.М. Сеченов выступил против идеалистической теории «свободы воли», характерной для реакционного мировоззрения.
Последние годы жизни Сеченов отдал изучению физиологических основ режима труда и отдыха человека. Он обнаружил много интересного, а главное, установил, что сон и отдых - разные вещи, что восьмичасовой сон обязателен, что рабочий день должен быть восьмичасовым. Но как физиолог, анализируя работу сердца, он пришел к выводу, что рабочий день должен быть еще короче.
3 Влияние трудов И.М. Сеченова на последующее развитие физиологии
Установив рефлекторную природу психической деятельности, Сеченов дал подробное толкование таких фундаментальных понятий психологии, как ощущения и восприятия, ассоциации, память, мышление, двигательные акты, развитие психики у детей. Впервые показал, что вся познавательная деятельность человека носит аналитико-синтетический характер психологического конгресса.
Опираясь на достижения физиологии органов чувств и исследования функций двигательного аппарата, Иван Михайлович подвергает критике агностицизм и развивает идеи о мышце, как органе достоверного познания пространственно-временных отношений вещей. Согласно Сеченову, чувственные сигналы, посылаемые работающей мышце, позволяют строить образы внешних предметов, а так же соотносить предметы между собой и тем самым служить телесной основой элементарных форм мышления.
Эти идеи о мышечной чувствительности стимулировали разработку современного учения о механизме чувственного восприятия, стали основой представления И.П.Павлова и его последователей о механизмах произвольных движений.
Огромной значение для развития отечественной нейрофизиологии имели такие работы И.М. Сеченова: «Физиология нервной системы) (1866) и особенно «Физиология нервных центров», в которых были обобщены и критически проанализированы как результаты собственных экспериментов, так и данные других исследований. Развиваемая в них идея о том, что регуляторная деятельность неровной системы осуществляется рефлекторно, надолго сделалась ведущей во всех исследованиях по физиологии центральной нервной системы.
И.М. Сеченов вооружил отечественную физиологию правильной методологией. Основным принципом Сеченова был последовательный материализм, стойкое убеждение, что в основе физиологических явлений лежит материальные физико-химические процессы. Второй принцип научной методологии И.М. Сеченова состоял в том, что изучение всех физиологических явлений должно осуществляться методом экспериментов. Электрофизиологические работы И.М. Сеченова способствовали распространению электрофизиологического метода для изучения физиологии нервов, мышц и нервной системы.
4 «Рефлексы головного мозга». Основной труд И.М. Сеченова
Весной 1862 г. Профессор Медико-хирургической академии Иван Михайлович Сеченов получил годовой отпуск и уехал за границу в Париж, где работал в лаборатории Клода Бернара. Здесь он делает открытие «центрального торможения рефлексов». И уже обдумывает основные положения будущего своего труда, получившего название «Рефлексы головного мозга».
Осень 1863 г. Сеченов выпускает статью по своей книге. Ученый отнес ее в «Современник». Первоначальное название статьи - «Попытка свести способы происхождения психических явлений на физиологические основы». В своем труде Сеченов утверждал, что вся разработанная психическая деятельность человека является ответом головного мозга на внешнее раздражение, причем концом любого психического акта будет сокращение тех или иных мышц.
Иван Михайлович был первым физиологом, который дерзнул начать изучение «душевной» деятельности теми же способами, какими изучалась деятельность «телесная», более того - первым, кто осмелился свести эту душевную деятельность к тем же законам, каким подчиняется телесная.
В редакции журнала «Современник» из-за цензурные соображений заглавие изменении: «Попытка ввести физиологические основы в психические процессы». Однако это не помогло. Свет по делам книгопечатания запретил печатать в «Современнике» работу Сеченова.
Несмотря на попытку властей скрыть работу Сеченова от общества, она очень скоро стала достоянием широких кругов читателей. О новых идеях всюду говорили, новые идеи обсуждали. Прогрессивная и мыслящая интеллигенция России зачитывалась Сеченовым.
Но власти рассудили по-иному. Они были насмерть перепуганы. Как «Отъявленный материалист», «идеолог нигилистов», профессор, состоящий под тайным надзором полиции, издает книгу. И власти приняли самые срочные меры, чтобы помешать автору поступить в более обширный оборот свое сочинение».
Дело было передано в Петербургский окружной суд «с покорнейшей просьбой о судебном преследовании автора и издателя книги «Рефлексы головного мозга» и об уничтожении самой книги».
В вину автору было поставлено то, что « Рефлексы головного мозга» якобы ниспровергают понятия о добре и зле, разрушают моральные основы общества. «Дело» попадает в прокуратуру судебной палаты, которая вынуждена признать, что «упомянутые сочинение проф. Сеченова не заключает в себе мыслей, за распространение коих сочинитель мог быть подлежащим ответственности». В свою очередь и министр внутренних дел вынужден был судебной преследование прекратить. 31 августа 1867 г. Книга была освобождена из-под ареста и поступила в продажу.
Иван Михайлович Сеченов приобрел в правительственных кругах репутацию «отъявленного материалиста», идеолога сил, враждебных устоям государства. Именно эта репутация помещала избранию его в адъютанты Академии наук, препятствовала утверждению его профессором Новороссийского университета.
Заключение
Своими работами И. М. Сеченов положил начало отечественной физиологии и создал материалистическую школу русских физиологов, которая сыграла важную роль в развитии физиологии, психологии и медицины не только в России, но и во всем мире. К. А. Тимирязев и И. П. Павлов называли И. М. Сеченова «гордостью русской мысли» и «отцом русской физиологии». Перефразируя слова Ньютона, сказанные о Декарте, можно утверждать, что Сеченов - самый крупный физиолог, на плечах которого стоит Павлов. «Честь создания настоящей большой русской физиологической школы и честь создания направления, определяющего в значительной степени развитие мировой физиологии, принадлежит Ивану Михайловичу Сеченову», писал выдающийся советский физиолог, академик Л. А. Орбели.
Сегодня очевидно, что многие современные разделы физиологии - нейрофизиология, физиология труда, спорта и отдыха, физико-химические (молекулярные) и биофизические направления в физиологии, эволюционная физиология, физиология высшей нервной деятельности, кибернетика и др. - своими корнями уходят к открытиям Ивана Михайловича Сеченова. Его работы составили в физиологии целую эпоху.
Список используемых источников
Анохин П.К. «От Декарта до Павлова».-М. : Медгиз, 1945М.Б. Мирский «И.М. Сеченов. Люди Науки.»
Березовский В.А. Иван Михайлович Сеченов. Киев, 1984;
Иван Михайлович Сеченов. К 150-летию со дня рождения / Под ред. П.Г. Костюка, С.Р. Микулинского, М.Г. Ярошевского. М., 1980.
Шикман А.П. Деятели отечественной истории. Биографический справочник. Москва, 1997 г.
Ярошевский М.Г. Иван Михайлович Сеченов (1829-1905). - Л.: Наука (Ленигр. отдел.), 1968
Батуев А.С. Высшая нервная деятельность. - М.: Высшая школа, 1991.
Батуев А.С., Соколова Л.В. К учению Сеченова о механизмах восприятия пространства.//Иван Михайлович Сеченов (К 150 летию со дня рождения) - М.:Наука, 1980.
Костюк П.Г. Сеченов и современная нейрофизиология.//Иван Михайлович Се-ченов (К 150 летию со дня рождения) - М.:Наука, 1980.
Черниговский В.Н. Проблема физиологии сенсорных систем в трудах Сеченова.//Иван Михайлович Сеченов (К 150 летию со дня рождения) - М.:Наука, 1980.сеченов физиология рефлекс
Сеченов И.М. Рефлексы головного мозга. - М.: Изд-во АН СССР, 1961.
Репетиторство
Нужна помощь по изучению какой-либы темы?
Наши специалисты проконсультируют или окажут репетиторские услуги по интересующей вас тематике.
Отправь заявку
с указанием темы прямо сейчас, чтобы узнать о возможности получения консультации.
Торможение - ослабление или прекращение какой-либо деятельности. Торможение в ЦНС открыл И.М.Семенов (1862 г.) в опытах на лягушках. Он сделал вывод, что в промежуточном мозге имеются центры, возбуждение которых тормозит двигательные центры спинного мозга.
Цель работы. Воспроизвести опыт И.М. Сеченова, доказывающий, что раздражение промежуточного мозга тормозит двигательные рефлексы.
Лягушка, препаровальные инструменты, штатив, булавки, скальпель глазной, марля, вата, 0,5%-ный р-р Н 2 SO 4 , кристаллики хлористого натрия, р-р Рингера, стакан с водой, 10%-ный этиловый спирт, чашка Петри.
Подготовка к работе. Лягушку наркотизируют р-ром этилового спирта, а затем заворачивают в марлевую салфетку (голова должна остаться свободной) и берут в левую руку, одновременно подпирая голову снизу указательным пальцем той же руки; малыми остроконечными ножницами, отступив от носовых отверстий к глазам, делают поперечный разрез кожи; надрезают кожу спереди назад по боковым поверхностям головы до места соединения головы с туловищем и отрезают кожный лоскут; ватой протирают обнажившиеся кости и ножницами делают поперечный разрез черепной коробки позади носовых отверстий; конец бранши малых остроконечных ножниц через поперечный разрез вводят под крышку черепа, прижимая острие изнутри к боковой поверхности крышки; по верхнебоковым поверхностям разрезают кости крышки до заднего конца полости черепа; освободившуюся костную пластинку приподнимают пинцетом и отрезают по заднему краю; обнажившуюся поверхность мозга осушают ватными шариками, перерезают мозг (рис. 7) и удаляют большие полушария.
Ход работы. Лягушку подвешивают на штативе и оставляют висеть, пока не кончится действие наркоза. Затем опыт проводят в следующей последовательности. К коже задней лапки прикладывают кусочек фильтровальной бумаги, смоченный р-ром Н 2 SO 4 , и определяют время рефлекса. После реакции лапку ополаскивают водой. Поверхность разреза мозга и черепную полость осушают ватными шариками и на зрительные бугры кладут кристаллик хлорида натрия. Через 1 мин к коже задней лапки вновь, прикладывают кусочек фильтровальной бумаги, смоченный р-ром Н 2 SO 4 , и определяют время рефлекса.
После ответной реакции ополаскивают лапку водой, удаляют кристаллик хлорида натрия и несколько раз поверхность мозга обмывают р-ром Рингера. Спустя 5, 10 и 15 минут опыт повторяют, раздражая заднюю лапку лягушки р-ром Н 2 SO 4 , в каждом случае определяя время рефлекса.
Работа 10. Наблюдение тонических рефлексов
Тонус скелетных мышц, необходимый для нормального положения тела в пространстве, обеспечивается тоническими рефлексами (статическими и статокинетическими). Центры тонических рефлексов находятся в продолговатом мозге (ядро Дейтерса) и в среднем мозге (красное ядро), а рецепторы - во внутреннем ухе (преддверие лабиринтов и полукружных каналов), мышцах и связках шеи, а также на коже.
Наибольшее значение имеют тонические рефлексы при изменении положения головы (перераспределение тонуса мышц шеи, туловища и конечностей), а также когда животное ложится и встает.
Цель работы . Наблюдать у животных рефлексы позы и выпрямительные рефлексы.
Объект исследования, материалы и оборудование. Лошадь, корова, небольшая собака, кролик, кошка, мягкая подстилка.
Ход работы.
Рефлексы позы.
1. Поднятие головы приводит к вытягиванию передних и сгибанию задних конечностей, а наклон головы дает противоположный эффект.
2. При повороте головы животного в сторону, конечность той стороны, куда повернута голова, выпрямляется, а противоположной - сгибается.
Выпрямительные рефлексы.
1. а) животное кладут спиной и теменем вниз (15 с удерживают в этом положении), затем освобождают голову - голова поворачивается теменем вверх (рефлекс с лабиринтов на шею ); б) передние конечности и плечевой пояс животного отпускают - передняя часть туловища и передние лапы поворачиваются в сторону поворота головы (рефлекс с шеи на туловище ); в) отпускают животное (нормальная поза восстанавливается).
2. Животное кладут на бок и удерживают голову в боковом положении, затем голову отпускают (она принимает нормальное положение теменем вверх), после этого животное отпускают (оно быстро вскакивает).
3. Выпрямительные рефлексы.
А. Кошку удерживают руками за лапы спиной вниз. С высоты 1-1,5 м животное отпускают. До приземления она переворачивается и встает на ноги. Отмечают, что в начале падения происходит поворот головы.
Б. Выпрямительные рефлексы у коровы и лошади появляются, когда они из положения лежа встают на ноги. Отмечают последовательные изменения положения головы и конечностей в процессе вставания.